
Hirundo 1/2024
Kanakulli saagi koostis ja selle muutused Eestis
Ülo Väli1,2, Jaan Grosberg1,2, Pelle Mellov1, Raul Melsas1, Rein Nellis1,3
1 Röövlinnutöörühm, Eesti Ornitoloogiaühing, Veski 4, 51005 Tartu
2 Elurikkuse ja loodusturismi õppetool, Põllumajandus- ja keskkonnainstituut, Eesti Maaülikool, Fr. R. Kreutzwaldi 5D, 51006 Tartu
3 Eluslooduse osakond, Keskkonnaagentuur, Mustamäe tee 33, 10616 Tallinn
Kokkuvõte
Kanakulli arvukus on Eestis viimastel aastakümnetel oluliselt kahanenud. Kirjanduse andmetel on selle liigi asurkonna langused olnud sageli seotud toidubaasi vähenemisega, kuid Eestis ei ole seda seost seni uuritud. Käesolevas töös koondasime aastatel 1993–2022 üle Eesti kogutud andmestiku (4963 saaklooma 237 pesapaigalt) kanakulli pesitsusaegse saagi koostisest, et analüüsida ajalisi muutusi erinevate saagirühmade osatähtsustes. Linnud hõlmasid 97,5% saakloomade arvust ning 98,6% biomassist, ülejäänud osa koosnes imetajatest (peamiselt oravatest). Olulisemateks saagirühmadeks olid vareslased (39,3% arvust ja 44,2% biomassist), tuvilised (23,3% ja 21,0%) ning kanalised (9,4% ja 15,0%), vähem tähtsad olid rästaslased (6,9% ja 1,6%), kurvitsalised (5,8% ja 4,1%), röövlinnud (4,7% ja 5,0%), partlased (3,9% ja 8,2%) ja rähnilised (1,9% ja 0,9%). Tähtsamateks saakliikideks olid hallvares, kodutuvi ning kaelustuvi, kes moodustasid kokku 48,6% saakobjektide arvust ning 55,5% massist. Peamiste saagirühmade osatähtsused muutusid viimase kolmekümne aasta vältel: vareslaste ja kodutuvi proportsioon vähenes, kuid kaelustuvi osa kasvas oluliselt. Viimastel kümnendite jooksul on kanakulli menüü muutunud mitmekesisemaks, mis sellel suhteliselt kitsale saagirühmale spetsialiseerunud liigil võib peegeldada toidupuudust. Tõenäoliselt on Eesti kanakulliasurkonna kahanemine olnud seotud erinevate linnurühmade (kanaliste, vareslaste) üheaegse arvukuse langusega või tuleneb hoopis muudest põhjustest, kuid selle kinnituseks on vajalikumad detailsemad uuringud. Samuti tuleks edaspidi analüüsida kanakulli talvise saagi koostist.
Sissejuhatus
Kanakull Accipiter gentilis on Eestis üldlevinud hajusalt pesitsev röövlind, kelle arvukus on viimase poolsajandi jooksul oluliselt vähenenud (Väli 2018, Elts et al. 2019). Möödunud sajandivahetusel dokumenteeriti kümne aasta jooksul koguni kahekordne kanakulli arvukuse langus (Lõhmus 1999, 2004, Väli et al. 2019), mistõttu seda liiki käsitleti ohualtina ning arvati II kaitsekategooriasse. Asurkonna kahanemise tõenäoliseks põhjuseks peeti Eesti maastikus aset leidnud suuri muutusi, eeskätt metsamajanduse intensiivistumist ning põllumaade sööti jäämist, mis mõjutasid saakloomade arvukust ja kättesaadavust (Lõhmus 1999, Väli 2018).
Kanakull on eeskätt puistutes saaki jahtima kohastunud haukaline ja tema peamiseks elupaigaks on erinevad metsad (Lõhmus 2001, Kenward 2006, Rutz et al. 2006). Plastilise käitumisega liigina on ta aga edukalt koloniseerinud ka mitmed teised elupaigad. Näiteks suudab kanakull edukalt saaki jahtida mosaiiksel kultuurmaastikul (Kenward 1982, Lõhmus 2001, Johansen et al. 2007, Mirski & Väli 2021) ning koguni linnades (Rutz 2006, 2008, Merling de Chapa et al. 2020). Kuna kanakull peab enamasti jahti kodupiirkonnas levinumatele sobivas suuruses saakobjektidele, peegeldub elupaigakasutuse varieeruvus ka saagi koostises (Kenward 2006, Rutz et al. 2006). Seega iseloomustab kanakulli saagi koostis üsna hästi elustiku seisundit ning kanakulli peetakse heaks keskkonnaindikaatoriks (Ozaki et al. 2006, Björklund et al. 2020, Natsukawa 2021). Teisest küljest võimaldab saagi analüüs mõista seoseid kanakulli elupaiga ning asurkonna seisundi vahel ning annab olulist informatsiooni tema kaitse korraldamiseks, liiati näitavad varasemad uuringud, et peamiste saakliikide asurkondade muutused peegelduvad ka kanakulli arvukuses (Linden & Wikman 1983, Rutz & Bijlsma 2006).
Eestis on kanakulli saagi koostis pälvinud tähelepanu erinevas kontekstis. Arvamus, et kanakull vähendab oluliselt nii ulukite kui kodulindude arvukust, oli peamiseks ajendiks 19. sajandi lõpus ja 20. sajandi esimesel poolel aset leidnud kanakulli ja teiste röövlindude tapmiskampaaniatele (Löwis 1898, Põllumees 1928, Kumari 1954, Kostrzewa 2008). Siiski jäid tolleaegsed kokkuvõtted saagi koostisest üldsõnalisteks (nt Põllumees 1928, Juhtund 1937). Alles 20. sajandi keskpaigas kogutud andmestiku põhjal (üle 200 murdmisjälje aastatelt 1954–1972) andis saagirühmade suhtelisest osatähtsusest ülevaate T. Randla (1976), kuid siis juba röövlindude kaitse kontekstis. Hiljem on aastate 1987–1992 saagiandmestikku põhjalikult analüüsinud A. Lõhmus (1993) ning 20. sajandi lõpu ja 21. sajandi alguse saagi koostisest tehti käsikirjaline kokkuvõte kanakulli kaitse tegevuskava tarbeks (Väli & Tuule 2012). Lisaks nendele üldistele uuringutele on meil saagi koostist uuritud ka piirkondlikult, näiteks Aakre ümbruses Valgamaal (Naaber 1965, Randla 1976) ja Härjanurme seirealal Tartumaal (Väli & Laansalu 2002).
Käesolevas töös koondasime viimase kolmekümne aasta jooksul üle Eesti kogutud mahuka andmestiku kanakulli saagi koostisest, et analüüsida ajalisi muutusi ning piirkondlikke erinevusi saagirühmade osatähtsustes. Meie hüpoteesiks oli see, et kanakulli arvukuse langus on olnud seotud teatud saakobjektide arvukuse kahanemisega, mis ühtlasi avaldub kanakulli saagi koostise muutustena. On ka võimalik, et saak on püsinud samasugusena, mis näitaks, et arvukuse langus võib olla seotud erinevate saagirühmade arvukuse üheaegse kahanemisega või põhjustatud hoopis mingi muu teguri poolt. Lisaks viimastel aastakümnetel toimunud muutuste jälgimisele võrdleme saadud tulemusi varem avaldatud kokkuvõtetega, et selgitada pikaajalisi muutusi Eesti kanakullide toitumises.
Materjal ja metoodika
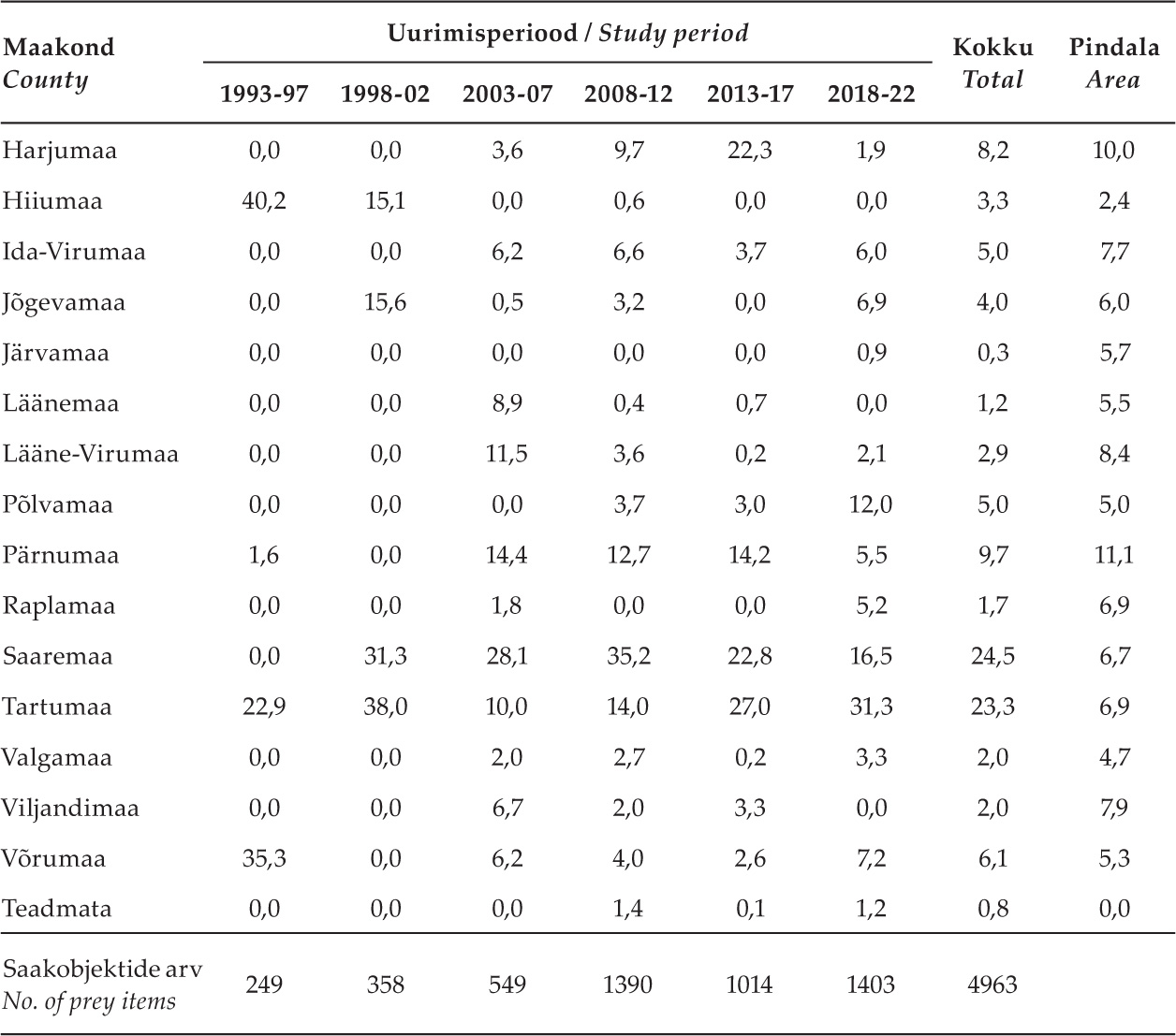
Aastatel 1993–2022 kogusime 4963 saakobjekti 237 kanakulli pesitsusterritooriumilt üle Eesti. Kogutud valim ei jaotunud Eesti erinevate piirkondade vahel siiski ühtlaselt. Maakondade võrdluses, võttes arvesse ka nende pindala, kogunes suhteliselt palju materjali Saare- ja Tartumaalt ning suhteliselt vähe Järva-, Lääne-, Lääne-Viru-, Rapla- ja Viljandimaalt (tabel 1). Ajaliselt jagasime andmestiku kuueks ajaperioodiks. Valimi maht oli esimestel perioodidel suhteliselt väike, kuid kasvas uuringu teisel poolel (tabel 1). Neid valimi omadusi võtame arvesse tulemuste interpreteerimisel.
Käesolevas töös uurisime kanakulli pesitsusaegset toitumist, analüüsides pesadest ning selle lähiümbrusest (pesaalune, mõnesaja meetri raadiuses asuvad saagipuhastuspaigad) kogutud saagijäänuseid. Seega näitavad käesoleva töö tulemused eeskätt poegadele toodava saagi koostist ning tähelepanu alt jääb välja vanalindude toitumine (v.a. emaslinnu poolt koos poegadega pesas söödav toit). Valdav enamik materjalist koguti poegade rõngastusajal, kuid põhjalikumalt uuritud maakondades (nt. Saaremaal, Tartumaal) otsiti saagijäänuseid pesade ümbrusest ka pesitsuse algusajal ning tehti spetsiaalseid kogumiskäike ka pärast pesitsust.
Saagi koostise analüüsimisel kasutasime nii luid, sulgi kui ka räppetompe. Säärane materjali kombineeriv analüüs annab suhteliselt hea ülevaate saagi koostisest (Lewis et al. 2004, García-Salgado et al. 2015). Siiski alahinnatakse isegi niisuguse analüüsiga linnupoegade osa, sest nendest ei jää enamasti pessa mingeid jäänuseid, samuti leitakse suhteliselt vähe väikeseid saakobjekte, kelle jäänused võivad kergesti kaduda pesamaterjali sekka (Sulkava 1964, Grønnesby & Nygard 2000). Alahinnatakse ka kõige suuremaid objekte, kelle kõiki osi ei pruugita pessa tuuagi (Rutz 2003) või kelle jäänused eemaldavad pesadest emaslinnud ning pesa alt kiskjad (nt rebane Vulpes vulpes, kährik Nyctereutes procyonoides) sagedamini kui väikeste saakloomade jäänused (Sulkava 1964). Röövlindude puhul tuleb ette pesade ristkasutust, näiteks võivad kanakullipesi pesitsemiseks kasutada ka händkakk Strix uralensis ja habekakk S. nebulosa ning hiireviu Buteo buteo, puhkepaigana ka metsnugis Martes martes. Seetõttu pole välistatud, et mõned saakloomad (näiteks pisiimetajad) võivad pärineda ka teistelt liikidelt. Siiski annab käesolevas töös kasutatud meetod seni parima ülevaate liigi pesitsusaegsest toitumisest kogu Eesti piires ning on piisav ühtmoodi uuritud ajaperioodide omavaheliseks võrdluseks.
Tabel 1. Kogutud saakloomade protsentuaalsed osatähtsused (arvu järgi) Eesti maakondades uurimisperioodide kaupa ning kogu uurimisaja jooksul kokku. Esitatud on ka iga maakonna pindala protsentuaalne osatähtsus Eesti maismaast (45 228 km2; enne 2017. aasta haldusreformi).
Table 1. Percentages of collected prey animals (by number) in Estonian counties by study period and in total during the entire research period. The percentage of the land area of each county in Estonia (45,228 km2; before the 2017 administrative reform) is also presented.
Saakloomad määrasime peamiselt autorite võrdluskogude ning vastava kirjanduse abil (Siivonen & Sulkava 1996, Brown et al. 2003, R. Tornberg, käsikiri), viimastel aastatel on eeskätt sulgede määramisel olnud abiks ka spetsiaalsed interneti-fotokogud (nt Haase et al. 2022, Schubert 2022). Kõiki liike ei olnud siiski alati võimalik eristada. Näiteks ei saanud enamasti vahet teha kodutuvil Columba livia ja õõnetuvil C. oenas ning hallvaresel Corvus corone cornix ja künnivaresel C. frugilegus, kuid kodutuvi ja hallvarese kohtamise tõenäosused on üleriigilises mastaabis oluliselt suuremad kui õõnetuvil ja künnivaresel. Samuti ei eristanud me üksnes luude materjalis leidumise korral sarnase suurusega vareslasi (hakk Corvus monedula ja harakas Pica pica) ning rästaliike (laulurästas Turdus philomelos ja vainurästas T. iliacus, hallrästas T. pilaris ja hoburästas T. viscivorus). Saakloomad jagasime 12 ökoloogilisse rühma: kanlised, kurvitsalised, partlased, röövlinnud (haukalised, pistrikulised, kakulised), tuvilised, rähnilised, vareslased, rästaslased, muud värvulised, muud linnud, imetajad, muud saakobjektid (teised klassid: kahepaiksed, roomajad). Seejuures kaasasime rästaslaste rühma ka kuldnoka (Sturnus vulgaris).
Kuna meie üheks eesmärgiks oli võrdlus varasemate töödega kanakulli saagist Eestis (Randla 1976, Lõhmus 1993), koondasime materjali eelmiste uuringutega sarnaselt kogu Eesti või piirkonna (maakonna) tasemel, mitte üksikute pesade kaupa. Seda lähenemist õigustas ka suhteliselt väike keskmine saakloomade arv ühe pesa pesitsusaasta kohta (11,3 ± 9,4 (SD) saakobjekti), mis tähendanuks pesapõhisel analüüsil märkimisväärset juhuslikkuse mõju ning suurt pesadevahelist saakloomade osatähtsuste varieerumist. Saakloomade arvu määrasime iga pesapaiga ja uurimisaasta kohta eraldi, loendades iga saakloomaliigi eri tüüpi luud või suled (hoo- või tüürsuled) ning võttes arvesse nende paarilisust. Osatähtsused arvutasime eraldi nii isendite arvu järgi kui ka kasutades nende keskmist kaalu, mille arvutasime Kumari (1954) ja Siivoneni ja Sulkava (1996) järgi. Saakloomade isendite arv iseloomustab eeskätt vanalindude panuse jaotumist saagijahil – iga saaklooma püüdmine nõuab ju iseseisvat pingutust (tõsi küll, suurema saaklooma surmamine ja transport nõuab vanalinnult rohkem energiat), saakloomade kaalu osatähtsused näitavad poegade üleskasvatamiseks vajaliku energiahulga jaotumist.
Kogu uuringu koond-osatähtsused esitame tekstis kuue ajaperioodi keskmisena, kuid võrdlevalt arvutasime osatähtsused ka kogu valimi põhjal, mis andis sarnase tulemuse (tabel 2). Osatähtsuste muutuse olulisust hindasime χ2-testiga, võrreldes iga saagirühma registreeritud isendite arvu jaotust antud rühma püsivale osatähtsusele vastava jaotusega, mis arvutati saagirühma osatähtsusest kogu uurimisperioodi valimis. Testide suure arvu tõttu korrigeerisime olulisuse tõenäosuse väärtusi Holm-Bonferroni meetodiga (Holm 1979).
Tulemused
Saagi koostis
Eesti kanakullide saak koosnes aastatel 1993–2022 valdavalt lindudest, kes moodustasid saakloomade arvust 97,5% ning biomassist 98,6%. Imetajad hõlmasid arvuliselt 2,4% ja massist 1,4%, teiste klasside osaks kokku oli arvuliselt 0,1% ja massist alla 0,1%. Lindudest olulisimaks seltsiks osutusid värvulised (47,9% ja 45,9%; siin ja edaspidi tähistab esimene number osatähtsust arvu järgi, teine number osatähtsust biomassi alusel), valdavalt vareslased (39,3% ja 44,2%) ning rästaslased (6,9% ja 1,6%). Suure osa hõlmasid ka tuvilised (23,3% ja 21,0%) ning kanalised (9,4% ja 15,0%). Vähem tähtsad olid kurvitsalised (5,8% ja 4,1%), röövlinnud (4,7% ja 5,0%), hanelised (valdavalt partlased: 3,9% ja 8,2%) ja rähnilised (1,9% ja 0,9%).
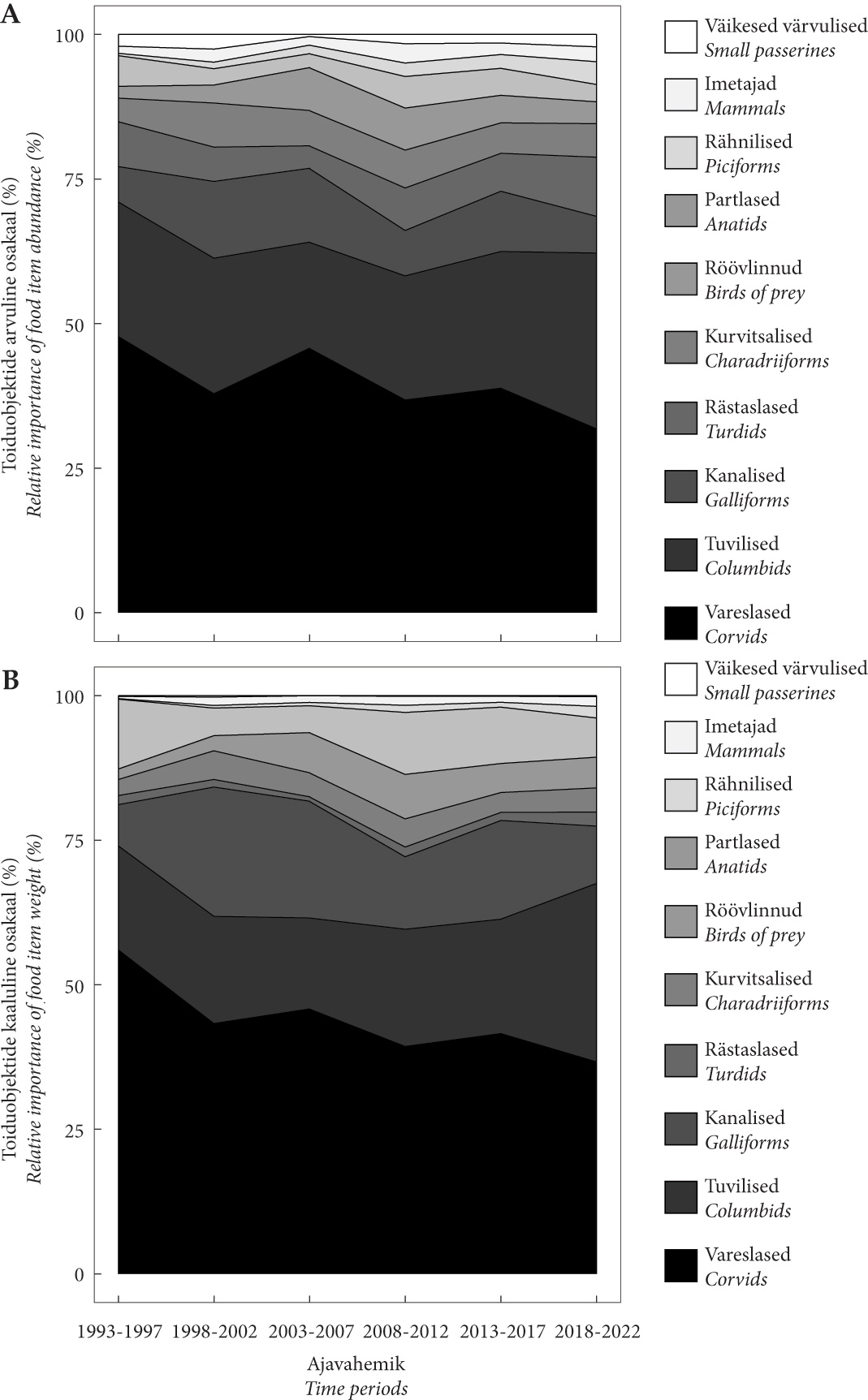
Joonis 1. Kanakulli saagi koostise muutused Eestis 1993–2022 hinnatuna saakloomade arvu (A) ja massi (B) järgi. Perioodide osatähtsuste täpsed väärtused on esitatud lisas 2.
Figure 1. Changes in the goshawk’s prey composition in Estonia 1993–2022, evaluated by the number (A) and mass (B) of prey animals. The exact values of the proportions of the periods are presented in Appendix 2.
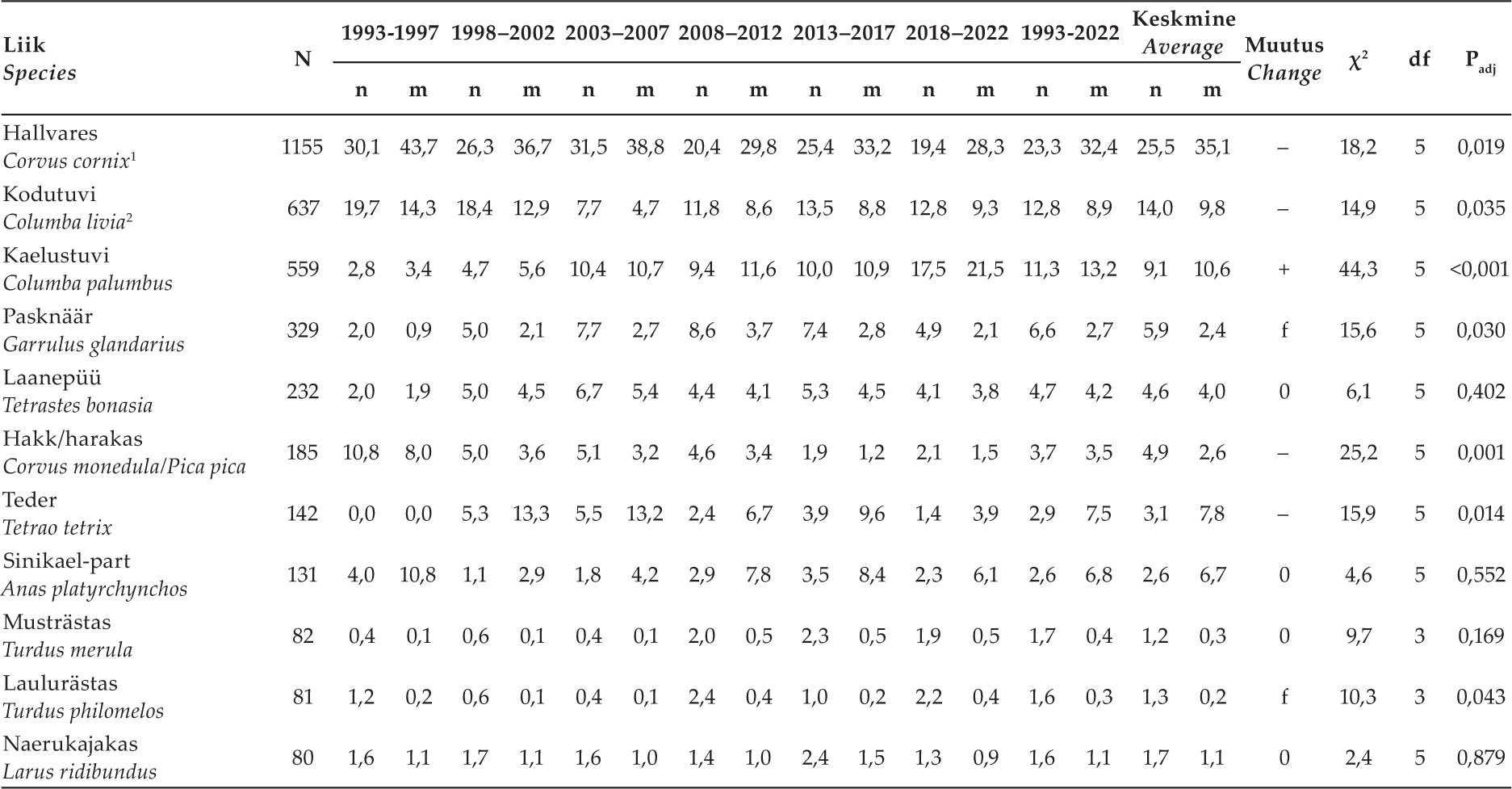
Kokku tegime kindlaks 99 liigi esinemise saagis (Lisa 1). Arvukamate liikide osatähtsused saakloomade koguarvust ja biomassist on esitatud tabelis 2. Tähtsamateks saakliikideks olid hallvares, kodutuvi ning kaelustuvi, kes moodustasid kokku 48,6% saakobjektide arvust ning 55,5% biomassist. Sageli murti ka teisi vareslasi (hakki, harakat ja pasknääri Garrulus glandarius). Kanalistest olid olulisemateks liikideks laanepüü Tetrastes bonasia ning teder Tetrao tetrix, harva murti ka nurmkanu Perdix perdix (n = 24; 1,1% ja 1,0%) ja metsiseid Tetrao urogallus (üksnes emas- ja noorlinde, 0,3% ja 1,3%). Kodukanu Gallus gallus domesticus leidsime vaid kolm isendit (0,1% ja 0,4%). Kurvitsalistest leidus enim naerukajakaid Larus ridibundus, metskurvitsaid Scolopax rusticola ning kiivitajaid Vanellus vanellus, partlastest sinikael-parte Anas platyrchynchos, vähemal määral ka piilparte A. crecca (0,6% ja 0,5%). Röövlindudest murti regulaarselt raudkulle Accipiter nisus ja kõrvukrätse Asio otus. Võrdlemisi sageli tabasid kanakullid ka suur-kirjurähne Dendrocopos major, vähem musträhne Dryocopus martius (0,7% ja 0,6%) ja kägusid Cuculus canorus (0,4% ja 0,1%). Imetajatest leidus kanakulli saagis regulaarselt vaid oravaid Sciurus vulgaris ning pisinärilisi (0,7% ja 0,03%).
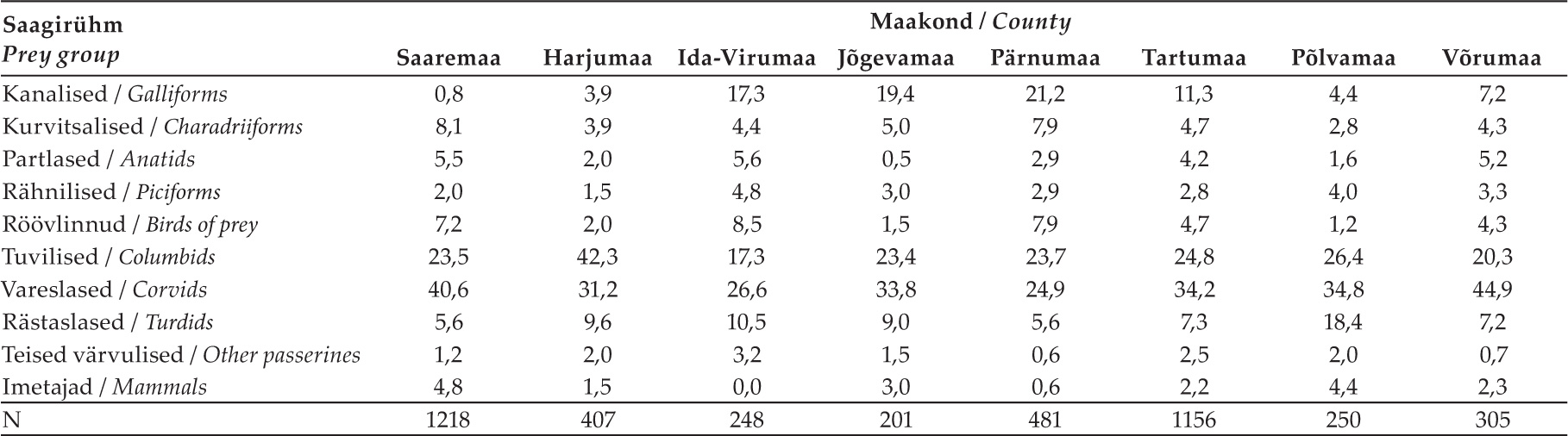
Kanakulli saagis esines piirkondlikke erinevusi (tabel 3). Vareslased olid kõigis paremini uuritud maakondades (valim suurem kui 200 saaklooma) arvukaimaks saagirühmaks, kuid eriti suure osakaaluga Saaremaa ja Võrumaa kanakullide saagis. Rästaslased olid sagedasimad Põlvamaal, tuvilised aga Harjumaal (sh Tallinn). Kanaliste osa oli suurim Pärnumaal, Jõgevamaal ja Ida-Virumaal, kurvitsalised Saaremaal, röövlinnud Ida-Virumaal, Pärnumaal ja Saaremaal.
Saagi koostise ajalised muutused
Arvulistest muutustest (joonis 1A, lisa 2) oli silmatorkavaim vareslaste osatähtsuse oluline kahanemine viimase kolmekümne aasta vältel (χ2 = 13,3; P= 0,041). Seejuures muutusid oluliselt nii hallvarese, pasknääri kui hakkide-harakate osatähtsused (tabel 2). Tuviliste (χ2 = 17,4; Padj = 0,012) osatähtsus kasvas viimasel perioodil. Kõigi perioodide lõikes oli märkimisväärne kaelustuvi Columba palumbus tõus, kuid samas langes kodutuvi osa (tabel 2). Ka rästaslaste osa kasvas viimasel perioodil (χ2 = 13,8; P= 0,043). Rähniliste osatähtsus tõusis ühtlaselt kogu uuringu vältel (χ2 = 11,2; P= 0,040; kahe esimese ajaperioodi väikesed valimid liideti), seejuures kasvasid nii suur-kirjurähni kui musträhni proportsioonid, ehkki need tõusud olid statistiliselt mitteolulised (tabel 2, lisa 2). Kanaliste (χ2 = 15,8; P = 0,037) ning röövlindude (χ2 = 15,8; P= 0,075) kõrgeimad osatähsused jäid keskmistele uurimisperioodidele, kuid näiteks tedre puhul võis näha pikaajalist olulist langust (tabel 2). Kurvitsaliste osatähtsuses muutust ei ilmnenud (χ2 = 2,6; P= 0,764).
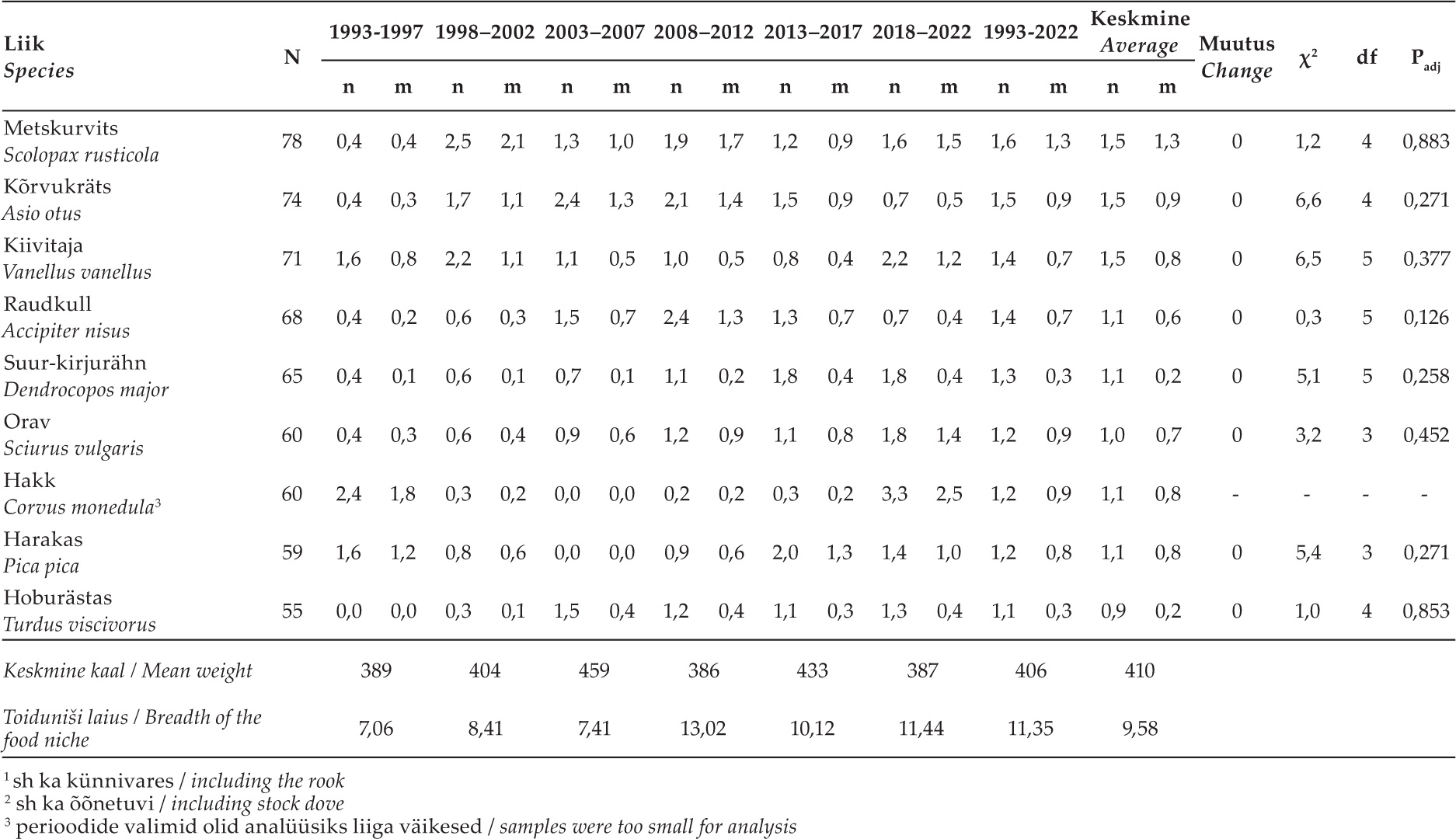
Tabel 2. Olulisemate saakloomade (üle 50 isendi) osatähtsuste, saakloomade keskmise kaalu ning toiduniši laiuse muutused 1993–2022. Iga saakobjekti puhul on esitatud isendite arv koguvalimis (N) ning protsentuaalsed osatähtsused arvu (n) ja kaalu (m) järgi, mis on arvutatud kõigi saakobjektide põhjal (vt lisa 1). Muutuste (+: tõus, –: langus, 0: stabiilne, f: fluktueeruv) olulisust hinnati χ2- testiga, esitatud on statistiku väärtus (χ2), vabadusastmete arv (df) ja korrigeeritud olulisuse tõenäosus (Padj).
Table 2. Changes in the proportions of the most important prey animals (more than 50 individuals), the average weight of prey animals, and the width of the food niche 1993–2022. For each prey object, the number of individuals in the total sample (N) and percentages by number (n) and weight (m), calculated from all prey objects, are presented (see Appendix 1). The significance of changes (+: increase, –: decrease, 0: stable, f: fluctuating) was evaluated with the χ2 test, the statistic value (χ2), the number of degrees of freedom (df) and the corrected significance probability (Padj) are presented.
Biomassi arvestades oli muutused samasugused, kuid neljal olulisemal saakrühmal veelgi ilmekamad (joonis 1B). Tartu- ja Saaremaal, kus materjali korjati ühtlaselt peaaegu terve uurimisperioodi vältel, võis täheldada ühesuguseid muutusi (Lisa 3).
Toidunišš oli uuringu teisel poolel märksa laiem kui esimesel poolel (tabel 2). Saakloomade keskmine kaal varieerus periooditi vahemikus 384–459 g, ilma kindla suundumuseta (tabel 2). Perioodide keskmine saaklooma kaal oli 410 ± 30 (SD) g.
Tabel 3. Saakrühmade arvulised osatähtsused paremini uuritud maakondades (n > 200).
Table 3. Proportions (% by numbers) of prey groups in best studied counties (n > 200).
Arutelu
Kanakulli saagi üldine koostis Eestis tänapäeval
Eesti kanakullide saagist ülekaaluka enamuse moodustavad linnud. Valdavalt lindudest toitumine on Euroopa kanakullidele tüüpiline ning iseäranis suur on lindude osa just Ida-Euroopas, kus imetajate osa ei küüni üle 10% (Rutz et al. 2006, Kenward 2006). Siiski on Eestis leitud lindude kõrge osatähtsus (ca 98%) isegi Ida-Euroopa ulatuses märkimisväärne. Tõenäoliselt on üheks põhjuseks sobivas suuruses imetajate vähesus. Optimaalses suuruses saakloomadest leidub meil vaid oravaid ning teda esineski ainsa imetajaliigina kanakulli saagis märkimisväärses koguses. Vähesel määral toitusid Eesti kanakullid ka pisinärilistest (eeskätt varakevadel) ning jänestest Lepus sp., kes on aga vastavalt märksa väiksemad või suuremad optimaalsest saagi suurusest (seda käsitleme põhjalikumalt allpool); pesitsusajal tabatud jänesed olid enamasti noorloomad. Mujal Euroopas on imetajatest lisaks oravatele suurim roll küülikutel (Oryctolagus cuniculus; Kenward 2006), keda Eestis ei leidu.
Lindudest moodustasid valdava enamiku saagist vareslased ja tuvilised. Vähemal määral, kuid siiski märkimisväärselt, leidus kanalisi, kurvitsalisi ja rästaslasi. Säärane saagi koostis on väga sarnane Lõuna-Soomele (Solonen et al. 2019). Kesk-Euroopas moodustavad tavaliselt peamise osa saagist tuvilised (eeskätt kodu- ja kaelustuvid), vareslased ja rästad (Rutz et al. 2006, Kosztrewa 2008). Soomes on aga oluline roll kanalistel. Seejuures leidub Põhja-Soome kanakullide saagis nii metsist, tetre, raba- ja laanepüüd (Tornberg & Sulkava 1991), kuid Lõuna-Soomes on olulisim osa laanepüül (Lindén & Wikman 1983), nii nagu Eestiski.
Leidsime, et ka Eesti-siseselt esinevad märkimisväärsed piirkondlikud erinevused. Need peegeldasid ilmselt maastikulisi iseärasusi, aga ka saakliikide levikut. Näiteks leidus rohke loodusmaastikuga Ida-Virumaal, Pärnumaal ja Jõgevamaal saagis suhteliselt arvukalt kanalisi ja röövlinde, kultuurmaarohkel Põlvamaal rästaslasi ning urbaniseerunud Harjumaal (sh Tallinnas) tuvilisi. Saaremaal, kus kanalisi on vähe (metsis ja laanepüü puuduvad sootuks), toitusid kanakullid olulisel määral vareslastest, aga suhteliselt sageli ka kurvitsalistest ning vee- ja röövlindudest. Niisiis määrab kanakulli saagi koostise piirkonna maastik, seda seost kavatseme detailsemalt analüüsida järgmistes uuringutes.
Eestis oli keskmine saakobjekti suurus 410 g. Meist põhja pool, Soomes, peetakse saagi optimaalseks suuruseks umbes 500 g (Tornberg & Sulkava 1990). Saksamaal kaalub enamik saakobjektidest 200–500 g, kuid keskmine kaal on vaid 255g (Engler et al. 2022). Lõuna-Euroopas eelistab kanakull saaki, mille kaal jääb vahemikku 100–400 g (Rebollo et al. 2017). Seega vastab Eestis saaklindude keskmine kaal, nagu saagi koostiski, meie geograafilisele asukohale ning on ilmselt seotud kullide endi suurusega, mis on Põhja-Euroopas suurem kui Lõuna-Euroopas (Kenward 2006). Üksikutel juhtudel langevad kanakulli saagiks ka tavapärasest suuremad linnud. Käesoleva töö andmestikus leidus vähesel määral, kuid siiski regulaarselt, emametsiseid, üksikisenditena esinesid valgepõsk-lagle Branta leucopsis, kalakotkas Pandion haliaeetus ja hallhaigur Ardea cinerea. Tõenäoliselt on need püüdnud emaslind, sest nii suured saakobjektid käivad isaslinnule üle jõu. Siiski on veebikaameras täheldatud koguni täiskasvanud must-toonekure Ciconia nigra ründamist isase kanakulli poolt, kuid sel juhul oli eesmärgiks ilmselt vanalinnu peletamine pesalt, et rünnata toonekure väikeseid pesapoegi (Kotkaklubi avaldamata andmed). Saagi kaal on suhteliselt suur kevadel, kui kanakullid jahivad täiskasvanud linde, ning see langeb suvel, kui looduses ning saagis leidub rohkem noorlinde (Toyne 1998).
Enamik käesoleva töö materjalist koguti rõngastusajal või pesitsuse järel, seega iseloomustavad meie tulemused eeskätt kanakulli toitumist pesitsuse keskel ja teisel poolel, ehk ajavahemikul maist juulini. See on periood, kui noorlinnud kasvavad kiiresti ning vajavad kõige enam toitu, mistõttu piisava saagi olemasolu on võtmetähtsusega poegade edukaks üleskasvatamiseks. Varasemate tööde põhjal iseloomustab seda perioodi suhteliselt suur vareslaste ja rästaslaste ning madalam kanaliste osa saagis (Lindén & Wikman 1983, Tornberg 1997). Selle eelistuse põhjuseks on rästaslaste ja vareslaste noorlindude lennuvõimestumine antud perioodil (Toyne 1998). Hiljem kasvab saagis kanaliste noorlindude osa (Lindén & Wikman 1983, Tornberg 1997). Soomes on leitud, et talvel on kanakulli saagis oluliselt suurem tähtsus imetajatel (Tornberg & Colpaert 2001). Sama võib oletada ka Eestis, kuid seni paraku vastavad uuringud puuduvad.
Pikaajalised muutused kanakulli toidus
Leidsime, et viimase kolmekümne aasta jooksul on kanakulli põhiliste toiduobjektide osatähtsused tema saagi koostises oluliselt muutunud. Esile võib tõsta vareslaste (eeskätt hallvarese) tähtsuse vähenemise ning tuviliste (eeskätt kaelustuvi) kasvu, kuid kahanenud on ka kanaliste proportsioon toidus. Järgnevalt käsitleme neid muutusi lähemalt ja võrdleme meie tulemusi varasemate töödega.
Vareslasi leidus möödunud sajandi keskel kanakulli saagis vaid 16% (Randla 1976), kuid 1990. aastate alguseks olid nad tõusnud olulisimaks saagirühmaks ja nende osatähtsus oli juba 27% (Lõhmus 1993). Möödunud sajandi lõpul ulatus vareslaste osa juba ligi pooleni saagist, kuid hiljem on vareslaste osatähtsus kanakulli saagis vähenenud, mis peegeldab ilmselt peamise saakliigi – hallvarese – arvukuse langust Eestis viimastel aastakümnetel (Elts et al. 2019), samamoodi on viimasel viieteistkümnel aastal kahanenud haraka arvukus (Elts et al. 2019). Ehkki metsavareslaste (pasknääri ja mänsaku Nucifraga caryocatactes) arvukus on viimastel aastakümnetel kasvanud, mis ilmneb ka nende osatähtsuse suurenemises kanakulli saagis, ei ole see muutus olnud piisav hallvarese kadumise kompenseerimiseks. Tõenäoliselt ei ole vareslaste osatähtsuse kahanemine seletatav valimi piirkondlike muutustega, sest selle rühma osatähtsus on kõrge kõigis paremini uuritud maakondades ning kahanemist võis täheldada nii Tartu- kui Saaremaal.
Möödunud sajandi keskel oli tuviliste arvuline osatähtsus kanakulli toidus vaid 15% (Randla 1976). 1990. aastate alguseks oli see kasvanud 19%-ni (Lõhmus 1993) ning käesoleva sajandi teise kümnendi keskpaigani püsiski tuviliste osa 18% ja 23% vahel. Viimastel aastatel on selle saakrühma roll märkimisväärselt kasvanud ja nüüd moodustavad tuvilised kanakulli saagist peaaegu kolmandiku (31%). Muutus ei ole seotud kanakulli „linnastumisega“, sest Tallinnast koguti märkimisväärses koguses saaki üksnes eelviimasel uurimisperioodil (2013–2017) kui tuviliste osatähtsuse tõus esile ei tõusnud. Liiati on kasvanud just asulatega vähem seotud kaelustuvi osatähtsus, mis viimasel uurimisperioodil hõlmas koguni ca 60% tuvide arvust ning ca 70% nende kogukaalust saagis. See peegeldab ilmselt kaelustuvi arvukuse pikaajalist kasvu (Elts et al. 2019).
Kanaliste osatähtsuse langus on seletatav selle rühma kõigi liikide arvukuse pikaajalise kahanemisega Eestis (Elts et al. 2019). Ka Põhja-Soomes on leitud otsene seos metsakanaliste arvukuse muutuste ja nende osakaalu vahel kanakulli menüüs (Kenward 2006, Tornberg et al. 2006). Aastatel 1998–2007 ilmnenud kanaliste osatähtsuse tõus kanakulli saagis ei tulenenud ilmselt kanaliste tegelikust tähtsuse kasvust, vaid pigem asjaolust, et sel perioodil koguti suhteliselt suur osa materjalist loodusmaastiku-rikastest maakondadest.
Põhiliste saakrühmade muutuste kõrval väärivad äramärkimist ka teiste saagirühmade osatähtsuste muutused, näiteks röövlindude, rähniliste ja imetajate (orava jt näriliste) kasvanud osa saagis. See näitab kanakulli menüü mitmekesistumist ning alternatiivide otsimist varasematele saakliikidele, mida kinnitab ka uuringu teisel poolel laienenud toidunišš. Nii menüü mitmekesistumine kui teiste röövlindude murdmine võib olla üheks indikaatoriks kanakulli toidupuudusest (Rutz & Bijlsma 2006). Röövlindude ja teiste suuremate saakobjektide sagedasem ründamine (näiteks on kanakullide ründeid üha rohkem täheldatud nii kalakotka, väike-konnakotka Aquila pomarina kui must-toonekure veebikaamerates) võib olla seotud ka kanakullide agressiivsuse kasvuga asurkonnas. Agressiivsed linnud, kes pesa juures isegi inimest ei pelga, lasti varem kullide tapmiskampaaniate ajal tõenäolisemalt maha kui ettevaatlikud isendid ning nende osatähtsus kahanes. Küllap on julgete ja agressiivsete lindude sagedus asurkonnas pikaajalise kaitse mõjul taas tõusnud, mis ilmneb ka hiljutises linnaasurkondade tekkes.
Kokkuvõtteks
Eesti kanakullide pesitsusaegne saak koosneb tänapäeval peamiselt vareslastest ning tuvilistest, kuid oma roll on ka teistel linnurühmadel. Imetajate osa on väga väike. Saagirühmade osatähtsused on viimase kolmekümne aasta jooksul mõnevõrra muutunud ja see on seotud peamiste saakliikide arvukuse muutustega. Käesoleva töö tulemused ei luba siiski tõmmata paralleele konkreetsete saakliikide või -rühmade muutuste ning möödunud sajandivahetusel täheldatud kanakulli arvukuse kahekordse langusega. On tõenäoline, et kanakulliasurkonna kahanemine oli seotud erinevate saagirühmade (kanaliste, vareslaste) üheaegse arvukuse muutustega, kuid selle kinnituseks on vajalikumad detailsemad uuringud. Loomulikult ei saa välistada ka teisi põhjuseid (näiteks mõnda haiguspuhangut), millele pole seni tähelepanu pööratud. Kindlasti tuleks edaspidi analüüsida kanakulli talvise saagi koostist, sest just talvine toit on kanakulli levila põhjaosas asurkonda oluliselt limiteeriv tegur (Tornberg et al. 2006).
Tänuavaldused
Oleme tänulikud kõigile röövlinnuhuvilistele, kes on aastakümnete jooksul otsinud ja seiranud kanakullide pesapaiku ning aidanud neist koguda materjali käesoleva töö analüüsideks. Määramisel olid meile abiks Urmas Abel, Tarmo Evestus, Arne Laansalu, Kristo Lauk, Triin Leetmaa, Asko Lõhmus, Renno Nellis, Gunnar Sein, Aarne Tuule jt.
Kasutatud kirjandus
- Björklund, H., Parkkinen, A., Hakkari, T., Heikkinen, R. K., Virkkala, R., Lensu, A. (2020). Predicting valuable forest habitats using an indicator species for biodiversity. Biological Conservation, 249, 108682.
- Brown, R., Ferguson, J., Lawrence, M., Lees, D. (2003). Tracks and signs of the birds of Britain & Europe. 2nd Edition. Christopher Helm, London.
- Elts, J., Leito, A., Leivits, M., Luigujõe, L., Nellis, R., Ots, M., Tammekänd, I., Väli, Ü. (2019) Eesti lindude staatus, pesitsusaegne ja talvine arvukus 2013–2017. Hirundo, 32, 1–39.
- Engler, M., van der Horst, Y., Merling de Chapa, M., Krone, O. (2021) How much to carry? Implications of maximum load carrying capacity for prey use of urban and rural Northern Goshawks Accipiter gentilis. Bird Study, 68, 433–442.
- García-Salgado, G., Rebollo, S., Pérez- Camacho, L., Martínez-Hesterkamp, S., Navarro, A., Fernández-Pereira, J. M. (2015) Evaluation of trail-cameras for analyzing the diet of nesting raptors using the Northern Goshawk as a model. PLoS One, 10, e0127585.
- Grønnesby, S., Nygard, T. (2000) Using time-lapse video monitoring to study prey selection by breeding goshawks Accipiter gentilis in central Norway. Ornis Fennica, 77, 117–129.
- Haase, A., Schlusen, J., Schwenk, K. (2022). featherbase.info – feather research and education. https://www.featherbase. info/
- Holm, S. (1979). A simple sequentially rejective multiple test procedure. Scandinavian Journal of Statistics, 6, 65–70.
- Johansen, H.M., Selas,V., Fagerland, K., Johnsen, J. T., Sveen, B., Tapia, L., Steen, R. (2007) Goshawk diet during the nestling period in farmland and forest-dominated areas in southern Norway. Ornis Fennica, 84, 181.
- Juhtund, N. (1937) Kulliliste toitumisest. Eesti Loodus, 1937, 219–220.
- Kenward, R. E. (1982) Goshawk hunting behaviour, and range size as a function of food and habitat availability. Journal of Animal Ecology, 51, 69-80.
- Kenward, R. E. (2006) The Goshawk. T & AD Poyser, London.
- Kumari, E. (1954) Eesti NSV linnud. Eesti Riiklik Kirjastus, Tallinn.
- Lewis, S. B., Fuller, M. R., Titus, K. (2004) A comparison of 3 methods for assessing raptor diet during the breeding season. Wildlife Society Bulletin, 32, 373-385.
- Lindén, H., Wikman, M. (1983) Goshawk predationontetraonids: availabilityofprey and diet of the predator in the breeding season. Journalof Animal Ecology, 51, 953-968. * Lõhmus, A. (1993) Kanakulli (Accipiter gentilis) toitumisest Eestis aastatel 1987–1992. Hirundo, 13, 3–14.
- Lõhmus, A. (1999) Röövlinnuprojekt aastatel 1994–1998. Hirundo, 12, 19–35.
- Lõhmus, A. (2001) Toitumisbiotoobi valikust Loode-Tartumaa röövlindudel. Hirundo, 14, 27 – 42.
- Lõhmus, A. (2004) Eesti röövlinnuseire 1999-2003: kanakulli kadu ja hiiretsüklite kellavärk. Hirundo, 17, 3–18.
- Kostrzewa, A. (2008) Nahrungswahl von Mäusebussard Buteo buteo und Habicht Accipiter gentilis – eine Metaanalyse rheinischer und europäischer Daten der letzten hundert Jahre. Charadrius, 44, 1-18.
- Löwis, O. V. (1898) Diebe und Räuber in der Baltischen Vogelwelt. J. Deubner, Riga.
- Mirski, P., Väli, Ü. (2021) Movements of birds of prey reveal the importance of tree lines, small woods and forest edges in agricultural landscapes. Landscape Ecology, 36, 1409–1421.
- Naager, J. (1965) Lahendamist vajav küsimus. Eesti Loodus, 1965, 312–315.
- Põllumees, J. A. (1928) Metsamehed, hävitage röövlinde ja loomi. Eesti Mets, 1928, 109 – 110.
- Randla, T. (1976) Eesti röövlinnud. Valgus, Tallinn.
- Rebollo, S., García-Salgado, G., Pérez- Camacho, L., Martínez-Hesterkamp, S., Navarro, A. Fernández-Pereira, J. M. (2017) Prey preferences and recent changes in diet of a breeding population of the Northern Goshawk Accipiter gentilis in Southwestern Europe. Bird Study, 64, 464–475.
- Rutz, C. (2003) Assessing the breeding season diet of goshawks Accipiter gentilis: biases of plucking analysis quantified by means of continuous radio-monitoring. Journal of Zoology, 259, 209–217.
- Rutz, C. (2006) Home range size, habitat use, activity patterns and hunting behaviour of urban-breeding Northern Goshawks Accipiter gentilis. Ardea, 94, 185.
- Rutz, C. (2008) The establishment of an urban bird population. Journal of Animal Ecology, 77, 1008–1019.
- Rutz, C., Bijlsma, R. G. (2006) Foodlimitation in a generalist predator. Proceedings of the Royal Society B: Biological Sciences, 273, 2069-2076.
- Rutz, C., Bijlsma, R.G., Marquiss, M., Kenward, R.E. (2006) Population limitation in the Northern Goshawk in Europe: a review with case studies. Studies in Avian Biology, 31, 158–197.
- Schubert, S. (2022) Vogelfedern – die Seite als Bestimmungshilfe für Mauserfedern und Rupfungen. http://vogelfedern.de/
- Solonen, T., Lokki, H., Sulkava, S. (2019) Diet and brood size in rural and urban Northern Goshawks Accipiter gentilis in southern Finland. Avian Biology Research, 12, 3-9.
- Sulkava, S. (1964) Zur Nahrungsbiologie des Habichts. Aquilo Ser. Zool., 3, 1–103
- Tornberg, R. (1997) Prey selection of the goshawk Accipiter gentilis during the breeding season: the role of prey profitability and vulnerability. Ornis Fennica, 74, 15-28.
- Tornberg, R., Colpaert, A. (2001) Survival, ranging, habitat choice and diet of the Northern Goshawk Accipiter gentilis during winter in Northern Finland. Ibis, 143, 41-50.
- Tornberg, R., Korpimaki, E., Byholm, P. (2006). Ecology of the northern goshawk in Fennoscandia. Studies in Avian Biology, 31, 141–157.
- Toyne, E. P. (1998). Breeding season diet of the Goshawk Accipiter gentilis in Wales. Ibis, 140, 569–579.
- Väli, Ü. (2018) Kanakull. Elts, J., Kuus, A., Leibak, E. (koost.) Eesti haudelindude levikuatlas, 290–291. Eesti Ornitoloogiaühing, Tartu.
- Väli, Ü., Nellis, R. Lõhmus, A. (2019) Eesti röövlindude pesitsusaegne arvukus ja sigimisedukus 1994–2018. Hirundo, 32, 40–62
- Väli, Ü., Tuule, A. (2012) Kanakulli tegevuskava eelnõu. Käsikiri.
Summary
The diet composition of the goshawk (Accipiter gentilis) and its changes in Estonia
The number of Goshawks has decreased significantly in Estonia in recent decades. According to the literature, declines in the population of this species have often been related to a decrease in the food base, but this connection has not been studied in Estonia so far. In the current study, we compiled a dataset (4,963 prey animals from 237 nest sites) collected across Estonia in the years 1993–2022 on the composition of the goshawk’s breeding season diet in order to analyse temporal changes in the importance of different prey groups. Birds comprised 97.5% of the number of prey and 98.6% of the biomass; the rest consisted of mammals (mainly squirrels). The most important prey groups were corvids (39.3% of the number and 44.2% of biomass), pigeons (23.3% and 21.0%), and grouse (9.4% and 15.0%); less important were thrushes (6.9% and 1.6%), charadriiforms (5.8% and 4.1%), birds of prey (4.7% and 5.0%), ducks (3.9% and 8.2%), and woodpeckers (1.9% and 0.9%). The most important prey species were the hooded crow, feral pigeon, and wood pigeon, which made up a total of 48.6% of the number of prey objects and 55.5% of the mass. The proportions of the main prey groups changed during the last thirty years: the proportion of corvids and the feral pigeons decreased, but the share of the wood pigeon increased significantly. In recent decades, the goshawk’s diet has become more diverse, which may reflect a lack of food for this species. It is likely that the decline of the Estonian goshawk population has been related to the simultaneous decline in the number of different groups of birds (grouse, corvids) or is due to other reasons, but more detailed studies are needed to confirm this. The composition of the goshawk’s winter diet should also be analysed in the future.
